An organism is any organic (Carbon-based) RNA/DNA protein which replicates & reproduces.
There is every indication that bacterial life has been present on the Earth for well more than three billion years. But more advanced forms of life have only erupted within the last billion, and most macroscopic animal forms have only appeared in the last half of that. This is because, in evolution, the biggest changes are those pertaining only to the smallest parts, the initial, fundamental, often microscopic structures. And once in place, these are also the hardest things to change. This is because DNA controls the organization of cells at two levels. At the most direct level, it controls the structure of the cell itself. In this capacity, the genetic code has already been fine-tuned for about a billion years longer than any multicellular organisms even existed. Because of this, the systems and mechanisms within the cell have been refined for extreme efficiency, and the genetic replication of the cell itself is almost perfect. However, the second level of replication is not in the construct of the cell, but in the assembly of numerous cells into complex (and wildly variable) multicellular configurations. The genetic code is significantly more prone to error in this expanded arena, especially since so many different arrangements will work. This means that the surface appearance of any organism may be dramatically different from its kin, but the foundational structures and mechanisms shared with them are much more difficult to change even over substantially longer periods of time. So there have been relatively few really significant macroevolutionary events since the dawn of life on this planet. Most of them were a really long time ago, and many of them have only been at the sub-cellular level, before there even were any multicellular anythings. Every other evolutionary stage is a comparatively trivial, inevitable consequence of an imperfect copying process amid variable environmental and reproductive pressures and endless functional possibilities.
To be an organism, one must also be organic. All life on Earth is Carbon-based. Carbon-based molecules are considered organic largely because some carbon-"chain" molecules were once only known to exist when they were created by living organisms. But the Carbon atom's special covalent-bonding properties indicate that, if there is life on other worlds, it may or may not have DNA. But it will likely still be Carbon-based. It will also almost certainly be dependant on water, again due to its unique covalent-bonding properties, and for the dual-polarity of that molecule.
"You, your joys and your sorrows, your memories and your ambitions, your sense of personal identity and free will,
are in fact no more than the behavior of a vast assembly of nerve cells and their associated molecules."
--Dr. Francis Crick; Nobel laureate, co-discoverer of the DNA molecule
One subset of organisms is Biota, living organisms, which are defined as any of the above
which can also maintain metabolic homestasis.
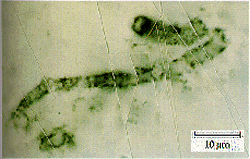
an apparently 3.5 bilion year-old
filamentous prokaryote microfossil
At its most basic, life is simply chemistry, and living tissues conform completely to those guidelines. The elements which form basic cell tissues for example create a phospholipid bilayer automatically upon contact with water, due to their combined polarity. Even the function of enzymes and transport vesicles and other miniscule but critical elements within a cell all conform to the functions of simple chemistry. And we know from the Urey/Miller experiment, that other organic compounds will also form automatically given the right chemical conditions. Consequently, there are a number of competing concepts to explain exactly how the first replicative polymers lead to the next stage, known as chemical hypercycles, and eventually on to protobionts, (quasi-living forms similar to viruses). Some of these hypotheses are considered scientifically credible, some are not. So far, we still don’t know which (if any) of the explanations posed so far is the right one. But if there’s one thing the wisdom of the ages has taught us, it is that simply not knowing the real reason is no reason to blame anything on magic.
“It is thus with regard to the disease [epilepsy] called Sacred:
it appears to me to be nowise more divine, nor more sacred than other diseases,
but has a natural cause from the originates like other affections.
Men regard its nature and cause as divine from ignorance and wonder, because it is not at all like to other diseases.
And this notion of its divinity is kept up by their inability to comprehend it...”
On Earth, everything we like to call "life" is also based on replicative RNA/DNA proteins. There are two main sub-categories of this, only one of which is actually considered to be alive, even though both can be killed by similar methods. Most biologists do not consider viruses to be alive because they lack metabolism. Metabolism means that cells can perform various chemical functions within themselves enabling them to maintain some level of homeostasis, a balanced internal environment. This again is due to the function of enzymes, chemical reactions, and molecular polarity. If there is any other aspect to life, science has yet to detect it. And if there is a supernatural componant to life, science will never be able to detect it.
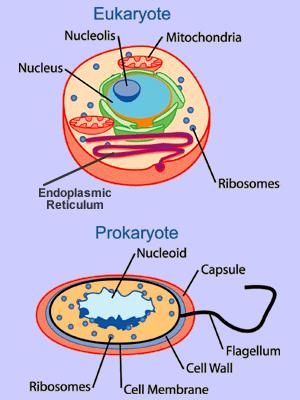
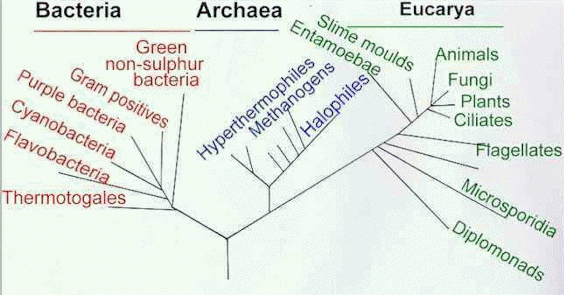
The most basic, (basal) fundamental, and encompassing sub-categories within "living organisms" are the domains, Eukarya and Prokarya. Prokarya is divided between Eubacteria and Archea, both often collectively described simply as bacteria. Prokaryotes are much smaller than Eukaryote cells, and do not contain a nucleus. Eukaryote cells do have a nucleus as well as some other sub-cellular features unique to this clade. Everything which produces eukaryote cells is defined as a Eukaryote itself. And with the exception of viruses, (which are not considered to be "true" life forms) everything that is undeniably alive is bacteria except for Eukaryotes. Or, from another perspective, the only living things that aren't eukaryotes are bacteria
If you are a eukaryote, and so are all the ancestors you ever knew of, -then obviously you have descended from eukaryotes. .
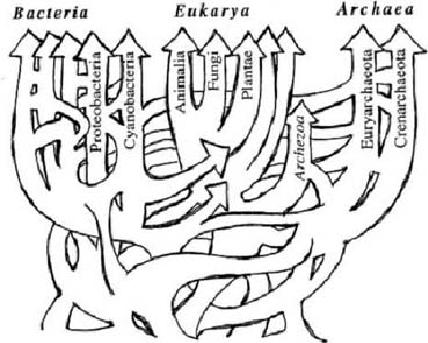
Now some would say we were all bacteria once. But that's not likely correct because eukaryotes technically did not 'evolve' from prokaryotes, though they evidently 'developed' from them some other way. 21st century revelations in genomic research implies there may be something more to the old idea of "primordial soup" then previously thought. There are now indications that at the root of each of the three main 'domains' of life, there was a sort of horozontal gene transfer going on which could not truly be considered part of the evolutionary process. By definition, evolution requires inherited genetic frequencies, but the requirement of "descent with modification" only allows for one series of ancestors rather than multiple lines of unrelated ones. While taxonomy still points to a single common ancestor for all eukaryotes, that ancestor seems to have had more than one mother! So at the point that true evolutionary common-ancestral phylogeny applies more or less exclusively to the domain of eukarya, they were already quite distinct of either of the "prokaryote" lineages.
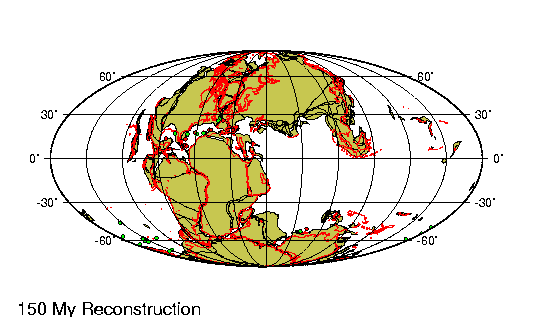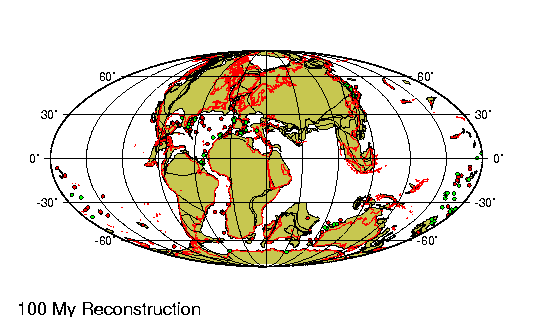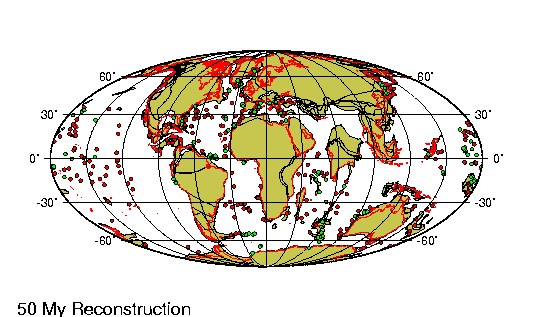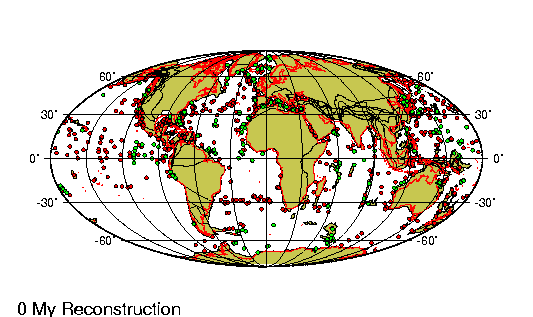
"The rooting of the Tree of Life, and the relationships of the major lineages, are controversial. The monophyly of Archaea is uncertain, and recent evidence for ancient lateral transfers of genes indicates that a highly complex model is needed to adequately represent the phylogenetic relationships among the major lineages of Life." --The Arizona Tree of Life project
One subset of life is Eukarya, metabolic organisms with nucleic cells.
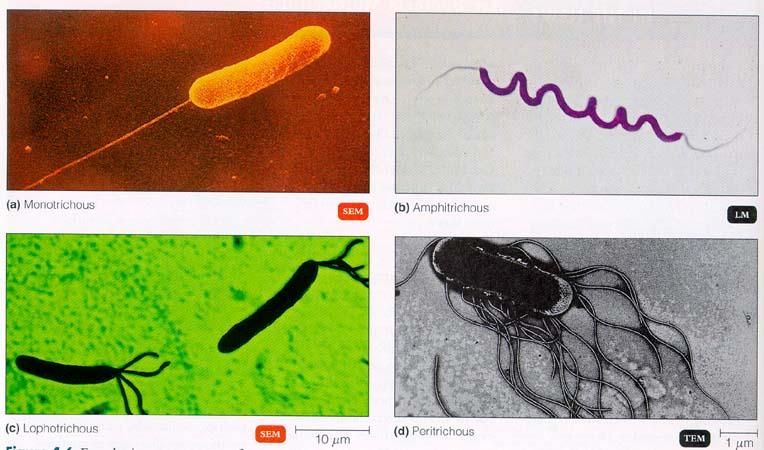
The domain, Eukarya consists of several sub-groups, including plants, algae, and those protists which appear to be collectively-related as opisthokonts. This opisthokont sub-category is not a formal taxon, but it is a unique grouping of organisms in which those cells that propel themselves with flagellum have that appendage mounted in the posterior "pushing" position. Most opisthokont cells don't have flagellum except on their reproductive spores or sperm. The flagellate cells of most other organisms either pull themselves with a front-mounted flagella, or they swim with myriad cilia.
One subset of Eukarya is Opisthokonta, Eukaryotes who's gammete cells have a single posterior flagellum.
Click here to add text.
Opisthokonts include microspores and their apparent descendants, fungi, as well as Choanoflagellates and all their apparent descendants in the animal kingdom. Now if all life were specially-created, you might think that some animals would have bikont reproductive cells, or sperm/spores which crawl on cillia like prokaryote cells do. But so far, not one animal or fungus is like that. They are all opisthokont, as if that trait appeared as an extremely rare and pivotal mutation among certain protists, and was then inherited by all the generations to flower out of that line since. The common ancestry model obviously explains this fact, but (to date) no would-be critic of evolution has ever been able to offer any alternative explanation for this or any of the other trends we see in taxonomy.
Opisthokonts are "nested" within the taxonomic kingdom, Protista. This is (I think) the only time that happens in the whole spectrum of taxonomy. Sister clades aren't usually nested in their own siblings. But the reason for it in this case is that Protists are such an ancient group that they're amazingly diverse, and have all been erroneously lumped into a single clade for ease of classification. Otherwise, there should be more than a dozen different kingdoms of life, and all but a very few of them would be Protists. Opisthokonts developed long before the Cambrian explosion, and even before the Vendian period. The bikont/opisthokont split evidently occurred in the Mesoproterozoic era, and represents one of the most significant developments in the history of life on Earth, second only to abiogenesis, and other pre-Camrbian transitions like the origin of sex, and the ascension to multicellular forms.
One subset of Opisthokonta is Metazoa, (Kingdom, Animalia)
multicellular opisthokonts which must ingest other organisms in a digestive tract in order to survive.
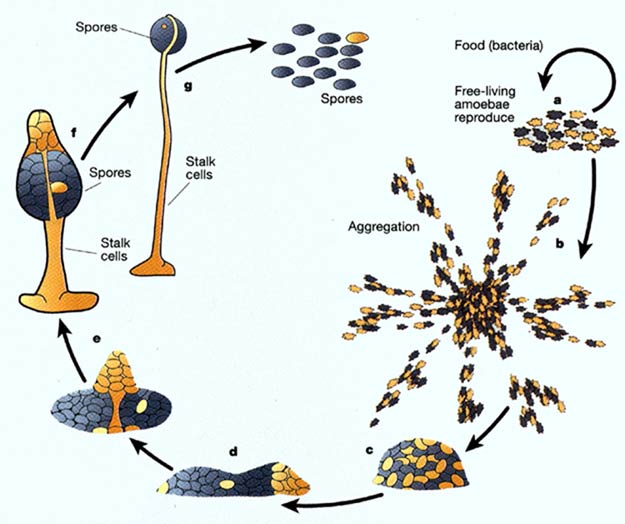
Those interested in transitional species should look at a group of protists called slime moulds, some of which appear to live as fungus and some as animals. The position of evolutionary theory and the model of common ancestry is that "simple" single-celled organisms are a lot harder to form than it is to build complex multicellular forms out of them. And there is substantial evidence in the record of microfossils that various forms of algae and such existed in different parts of the world's oceans as much as a billion years before anything recognizable as a plant or animal.
The slime mould, Dictyostelium discoideum represents the method by which scientist consider that life made the move from single cells to many cooperative cells, and is singularly representative of the most profound of all evolutionary stages. This is a communal organism, meaning that millions of individual cells amass themselves to function as a single entity. It travels as a sort of worm. But since it is made of amoebic cells, each cell has to nourish itself independently. So when it encounters food, the worm-unit dissolves itself into its cellular components to osmosize food before reassembling itself, and moving on. At some point, it seems clear that communal organisms like this stopped dissolving themselves every time they wanted to eat, once the movement of internal cells facilitated some means of internal distribution. These cells of course eventually became dependant on the surrounding body, and lost the ability to survive independent of it. Dead cells and waste material, (including calcium and keratin) was of course pushed to the outside where they became a covering, like skin, bone or shell, (in much later forms). In one particular Opisthokont sub-set, Metazoa, (animals) these hard formations appear to have begun as crystalline spicules. So those interested in finding transitional forms here should also look at choanoflagellates as an intermediate form from fungus-like protists to the mother of all animals.
Another of the evident trends of evolution is that the offspring of two closely-related organisms will be more primitive than its parents, in that it will look more like the common ancestor of both than the adults of either species will. And choanoflagellates very closely-resemble the nymph stage of sponges. Recently, a team of taxonomists, geneticists, and microbiologists working on the Shape of Life project at the Sogin Marine Biological Laboratory in Woods Hole, Massachusetts, -sampled DNA from dozens of animals, and from them, identified and sequenced a single gene common to every animal species known to man. This revealed a parent pattern, a genetic root fingerprint implicating the sponge as the most basal of all macroscopic animals.
All living organisms fall within two (or three) groups including Eukaryotes. The Domain, Eukarya consists of several subgroups including kingdom; Animalia. Animals are metazoan (multicellular). And what sets them apart from all other life forms is that they are unable to make their own food, and must ingest and digest other organisms in order to sustain themselves. Fungi do this too, but animals do it by way of an internal digestive tract. Animals are also identified at the cellular level by thier lack of cell walls, and they are always diploid except as gametes.
Animal: 'a-n&-m&l
1 : any of a kingdom (Animalia) of living things including many-celled organisms and often many of the single-celled ones (as protozoan) that typically differ from plants in having cells without cellulose walls, in lacking chlorophyll and the capacity for photosynthesis, in requiring more complex food materials (as proteins), in being organized to a greater degree of complexity, and in having the capacity for spontaneous movement and rapid motor responses to stimulation
--Merriam Webster Online Dictionary
animal (n-ml) n.
any organic (Carbon-based) replicative RNA/DNA protein organism:
(a) consisting of multiple diploid cells which each contain a nucleus;
(b) which perform chemical reactions and acheive homeostasis;
(c) who's gammete cells have a posterior flagella;
(d) which must ingest and digest other organisms in a digestive tract in order to sustain themselves.
--Biological definition
"I said in mine heart concerning the estate of the sons of men, that God might manifest them,
and that they might see that they themselves are beasts.
For that which befalleth the sons of men befalleth beasts;
even one thing befalleth them: as the one dieth, so dieth the other; yea, they have all one breath;
so that a man hath no preeminence above a beast: for all is vanity.
All go unto one place; all are of the dust, and all turn to dust again."
--Ecclesiastes; 3:18-20
Obviously, if you and all your ancestors that you know of are (by definition) animals,
then there shouldn't be any argument about whether you descended from animals.
Nor could you generalize differences between "humans and animals" for the same reason.
Instead, you could only cite differences between humans and certain other animals.
One subset of Metazoa is Eumetazoa,
which includes all animals more advanced than sponges.
We must understand that taxonomy is based as much on an organism's reproduction, life cycle and development as it is on the physiognomy or function of the form itself. The next few sub categories in this sequence should illustrate this.
For example, sponges aren't always sedentary. Their nymph form is mobile and transports itself to the site where it will settle down for the rest of its life. Some of the other primitive animal forms also have a nymph stage nearly identical to that of the sponge, particularly in the case of the hydra, which never seems to develop beyond that point. We are not actually descended from sponges, but from these sorts of nymphs, some of whom lead to sponges, while others lead to more advanced animals.
Corals and anemonies are also similar in this respect, and considered closely-related. Others may be similar to the sponge nymph too except that they remain mobile their entire lives. Comb jellies are like that, except a bit more sleek. And the last (and most advanced) group are slightly different in that they only represent the earliest phase of development, and begin life with a shape usually more like that of worms. Some of these can develop into beings that are much much more complex.
Sponges, the most basal of all animals, are classified in the sub-kingdom, Porifera. All other animals are in a sister subkingdom called Eumetazoa. Most of these develop differently than sponges in adulthood so that their cells are organized into various tissues. And except for the shapeless placozoans, they also have some degree of symmetry in that they are at least tubular with a definite front and rear. These include the cnidarians, the first animals to reveal the continuously repeating rhythmic pulse which scientists believe led to a heartbeat once such organs began to form.
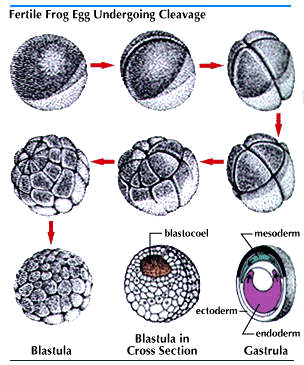
The first hints of the circulatory system appear in one line of a group of Eumetazoans which have achieved a bilateral symmetry. Almost all of the animals which come to mind, (including starfish) are bilaterally symmetrical; they can all be divided equally to provide mirror left & right images. But Bilateria has more criteria than just that. All bilaterally-symmetrical animals are triploblastic, which means they develop three germ layers;
The three germ layers are;
Ectoderm - Covers the surface of the embryo and forms the outer covering of the animal and also the central nervous system in some phyla.
Endoderm - the innermost germ layer which lines the archenteron (primitive gut).
It forms lining of the digestive tract, and outpocketing give rise to the liver and lungs of vertebrates.
Mesoderm - located between the ectoderm and endoderm. Forms the muscles and most organs located between the digestive tract and outer covering of the animal. The circulatory and (in vertebrates) the skeletal system stems from this
Note that only Bilateria have a mesoderm.
One subset of Eumetazoa is Bilateria, triploblast animals which at some stage of development are bilaterally-symetrical.
One subset of Bilateria is Coelomata, bilaterally-symetrical animals with an tubular internal digestive cavity.
There are two subdivisions of Bilateria, one based on body cavity or the lack thereof. Acoelomata have no body cavity present between the digestive tract and the outer body wall. Platyhelminthes (flatworms) belong here. All other animals have some sort of body cavity which joins a mouth of some kind at one end and an anus at the other.
In the Psuedocoelomata, a fluid-filled body cavity separates the digestive tract and the outer body wall. This cavity ( the psuedocoelom) is not lined, (or not completely lined) with mesodermal tissue. The so-called "Aschelminthes" (including the Nematoda, Rotifera and other groups of mostly tiny animals) are psuedocoelomate.
The true Coelomata have a body cavity which is completely tissue-lined, resulting in a "tube-within-a-tube" design. The fluid-filled body cavity completely lined with mesoderm, (the coelom) separates the digestive tract from the outer body-wall. Mesenteries connect the inner and outer mesoderm layers and suspend the internal organs in the coelom. All vertebrates and most invertebrates [arthropods] are coelomates, another Pre-Cambrian development.
The adaptive advantages of a body cavity are that it cushions the organs, preventing internal injury, and allowing them to grow independent of body wall. The non-compressible internal fluid also acts as a hydrostatic skeleton. [source; Paleos.com]
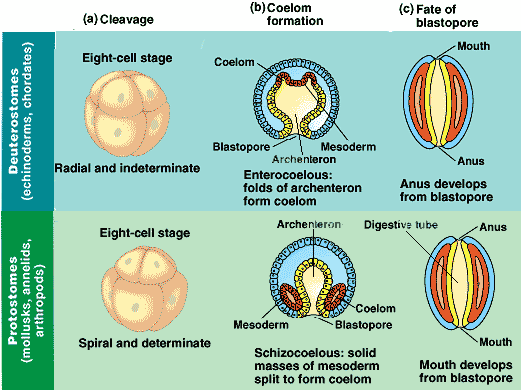
One subset of Coelomata is Deuterostomia, coelomates in which early development of the
digestive tract begins with a blastopore where the anal orafice opens before the oral one.
There are some bilateral animals that may not look bilateral, as they seem more like mosses or plants. But their bodies can still be divided left-to-right with equal sides. Most bilateral animals look like slugs, cucumbers, or worms. One group typically have multiple legs like caterpillars, and within that group are some with hard shells of keratin forming exoskeletons. These are the arthropods; insects, crustaceans, arachnids, and trilobites, etc. These also have rear-abdominal hearts which (again) appear to have been inherited from an ancestor shared in common.
Now this is a strange thing to have in common with every single 'higher" life form. All other life, arthropods, mollusks, bryozoans, and brachiopods, (etc.) all develop a connection from the blastopore to the mouth first. This is without exception in any case we've seen so far. If they were specially-created, one might think that any of them could develop by some other means, or in some other order. Maybe snails would develop like mammals, and fish develop like squids, something like that, something that wouldn't indicate an inherited trait consistent with both the genetics and morphology of common ancestry. But that is never what we see. Everything in nature consistently adheres to everything we would expect of a chain of inherited variations carried down through flowering lines of descent. Starfish, urchins, acorn worms and every single thing that ever had a spinal chord all develop the opening for the anus first. Isn't that odd?
All the various types of bilateral animals still fall within two basic types, which are each defined by a particular stage of their development. In all these animals, the blastopore, (the cavity that will become the digestive tract) develops internally, and eventually that internal cavity connects with an opening at one end of the animal, and then the other, completing the oral-to-anal passageway. In lophotrochozoans, ecdysozoans, and others, the mouth opens first, and the anus after that. But one line of bilateral animals got it backwards. In deuterostomes, the anus is the first to open the connection to the blastopore, and the mouth makes the connection after that.
Now, its not so strange that some members of these three groups, (which seem so different) have this one peculiarity in common, even though their genetics do indicate their relationship. But what is significant is that absolutely every last one of the [presumed] descendants of these basal forms (tested to date) share this developmental peculiarity too. That's one of many indications of common ancestry so far.
But wait, there's more!
One subset of Deuterostomia is Chordata, Deuterostomes with a spinal chord.
Evolution began only after another biological process
Critics of evolution often talk about the Cambrian "explosion" in an attempt to imply that that all the diversity of life and all the phyla known today appeared simultaneously at that time. But all the variation mentioned up to this point, occured prior to the Cambrian era, and all remaining developments yet to be discussed from this point in the sequence occur during or after the Cambrian era.
One specific line of Deuterostomes is evidently very closely-related to another by virtue of two factors, both possess a single central nervous structure and even pharyngeal gill slits. However, one of these is a worm! This worm with a mouth, anus, heart and gills, (which aren't typical of any other worm) also has a sort of spinal chord almost like that of a real (fully-developed) spinal chord. The family name for these worms is Hemichordata, which means "half-chordate". Those looking for transitional species here need look no further.
Every single modern chordate we know of so far appears to be a deuterostome at the same time. So it is reasonable to assume that Pikaia developed that way too, and lead to other invertebrate fish. The fossil record reveals that some of these spineless fossil fish also developed partially-calcified internal structures which lead to skeletons.
But the fossil record bears still more examples for the origin of chordates from creatures like this. Tiny swimming worms have been identified from a particular cache of Cambrian and Ordovician fossils in Canada's Burgess Shale. Some of these "worms" have teeth, (pretty serious ones too) even though they as yet have no jaws nor bone on which to mount jaws. One particular specimen called Pikaia is considered to be the oldest example of a true chordate. It is a sort of slender slug with a elaborate body-length fin for swimming, and a definite indication of a spinal chord, even though it didn't yet have a spine to keep it in.
One subset of Chordata is Craniata, Chordates with a brain enclosed in a skull.
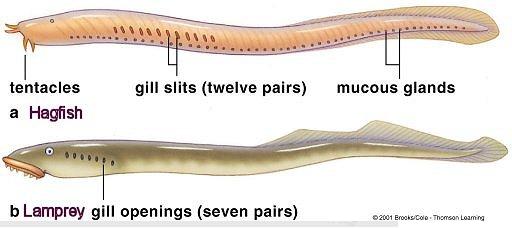
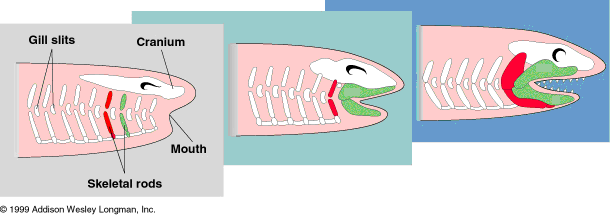
Not all chordates have a skull. Those that do are in the subphylum, Craniata. Hagfish are craniates in that they have a skull, but they have no jaws or spinal vertebrae to go with it. The lamprey has a skull, and vertebrae, but no jaws. Nothing has jaws without a skull or vertebrae, and that includes Chondrichthyes, (sharks and rays) where these things are there, but they're now made entirely of cartilage instead of bone, (except for the jaw itself). So it appears that the skull developed first, followed by vertebrae, and then the jaw. And this is consistent with what we see in the fossil record also, where the first fish to have skulls and backbones and all that still lacked jaws.
One subset of Craniata is Vertebrata, Craniates with a spinal vertebrae descending from the skull.
Not all craniates have vertebrae, living and fossil hagfish for example, (as well as other fossil fish) only have a skull with no spine either of cartilage or bone. Both of these materials form deposits in the body as an animal matures. Sometimes this presents a hindrance, like calcium deposits forming in human knee joints. But they can be advantageous too, like the way they evidently developed over time to form around the spinal chord in jointed segments allowing movement with additional strength. As with every other aspect of evolution, we have computer and robotic models to explain how it likely occurred. And as one might expect, the fossil record also reveals the earliest, (and definitely most primitive) transitional skeletal vertebrates from the Cambrian and Ordovician periods. But we have plenty of not-yet vertebrate fish contemporary to, and sometimes even older than these, again implying ancestry.
One subset of Vertebrata is Gnathostomata, vertebrates with a jawbone.
Acanthodians are the first fish known to have jaws. These placoderms appeared in the early Silurian, near the very dawn of vertebrate evolution. There were no other craniate fish that I know of to precede them except pre-gnathostomes like Haikouichthys ercaicunensis, a Cambrian fish, basal to anaspids and very close to the lampreys and Jamoytius, just as evolutionary theory predicts. Early Devonian Palaeoniscoids are thought to be descended from acanthodians as they still wore acanthodian scales. But bore a reduction in the placoderm traits such as the thickness of the head-shield. But the early Silurian appearance of Acanthodians was relatively early, and there doesn't appear to have been any other, more advanced [gnathostome] fish around at that time.
As you can see, the development of the jaw is in concert with the post-vertebrate expansion of the skull, and of course this is consistent with what we see in extant or extinct example species representative of this lineage, both among Chondrychthys and bony fish as well as their jawless, lamprey-like predecessors.
The first gnathostomes were the common ancestors of cartilaginous fish and vertebrates with more calcified bones. And we have some interesting composite fossils from that era that show traits inherited by a number of subgroups at once. For instance, compare those vicious bottom teeth to the rolling row system even some ancient sharks used for their teeth. This is where we find the common ancestors and transitional species leading from the earliest, and most primitive bony fish, to the earliest and most primitive ancestors of sharks and rays.
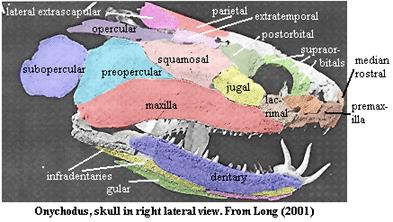
The skull at left, described as Onychodus, is misclassified. This is actually the skull of Quebecius quebecensis, (below) a predatory porolepiform lungfish from 360 million years ago..
"Psarolepis shares a number of characteristics previously believed to be unique to actinopterygians or sarcopterygians. In addition, it has several features, such as the fin spines and characters of the shoulder plate, which are associated with placoderms, chondrichthyans, or acanthodians. In overall appearance, Psarolepis most resembles a sarcopterygian, but the plates in isolation look strikingly like placoderm material."